Difference between revisions of "Diffusion of C and Ce"
(→Rate equation) |
(→Parameters) |
||
| Line 22: | Line 22: | ||
== Parameters == | == Parameters == | ||
| − | The parameter of this reaction is the diffusion rate of C (<math>D</math>). | + | The parameter of this reaction is the diffusion rate of C (<math>D</math>). In bacteria communication, the autoinducer can either passively diffuse in and out of the cell or, in cases of larger signalling molecules, can be actively transported through the cell membrane. Generally, the diffusion rates when driven by active processes are four orders of magnitude smaller than the passive diffusion rates of small molecules <ref name="Weber2011"> [http://www.biomedcentral.com/content/pdf/1752-0509-5-11.pdf Weber M., Buceta J. ''Noise regulation by quorum sensing in low mRNA copy number systems.'' BMC Systems Biology 2011, 5:11]</ref>. In this model, active transport is not explored, as SCBs are small molecules that can easily diffuse though the cell membrane. Moreover, transport driven by SCB concentration differences due to increase of the cell density in the environment, is simulated. This means that as cell density grows, more autoinducer molecules enter the cell from the outside. The parameter values were derived from measurements and estimations of different autoinducers' passive diffusion and active transport rates in bacteria. |
| − | {|class="wikitable" | + | {|class="wikitable" |
! Name | ! Name | ||
! Value | ! Value | ||
! Units | ! Units | ||
| − | ! | + | ! Value in previous GBL models <ref name="Mehra2008"> [http://www.plosone.org/article/fetchObject.action?uri=info:doi/10.1371/journal.pone.0002724&representation=PDF S. Mehra, S. Charaniya, E. Takano, and W.-S. Hu. ''A bistable gene switch for antibiotic biosynthesis: The butyrolactone regulon in streptomyces coelicolor.'' PLoS ONE, 3(7), 2008.] </ref> <ref name="Chatterjee2011"> [http://www.plosone.org/article/fetchObject.action?uri=info:doi/10.1371/journal.pone.0021974&representation=PDF A. Chatterjee, L. Drews, S. Mehra, E. Takano, Y.N. Kaznessis, and W.-S. Hu. ''Convergent transcription in the butyrolactone regulon in streptomyces coelicolor confers a bistable genetic switch for antibiotic biosynthesis.'' PLoS ONE, 6(7), 2011.] </ref> |
| − | ! Remarks | + | ! Remarks-Reference |
|- | |- | ||
|<math>D</math> | |<math>D</math> | ||
| − | |<math>0.1-24 </math> <ref name="Kaplan1985"> Kaplan HB, Greenberg EP. ''Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system''. Journal of Bacteriology. 1985;163(3):1210-1214.</ref> <ref name="Goryachev2005"> Goryachev AB, Toh D-J, Wee KB, Lee T, Zhang H-B, Zhang L-H (2005) ''Transition to Quorum Sensing in an Agrobacterium Population: A Stochastic Model.'' PLoS Comput Biol 1(4): e37.</ref> <ref name="Groisman2005"> A. Groisman, C. Lobo, H. Cho, J K. Campbell, Y. S. Dufour, A. M Stevens & A. Levchenko. ''A microfluidic chemostat for experiments with bacterial and yeast cells.'' Nature Methods 2005(2), 685 - 689 </ref> <ref name=" | + | |<math>0.1-24</math> <ref name="Kaplan1985"> [http://www.ncbi.nlm.nih.gov/pmc/articles/PMC219261/pdf/jbacter00220-0404.pdf Kaplan HB, Greenberg EP. ''Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system''. Journal of Bacteriology. 1985;163(3):1210-1214.]</ref> <ref name="Goryachev2005"> [http://www.ploscompbiol.org/article/fetchObject.action?uri=info:doi/10.1371/journal.pcbi.0010037&representation=PDF Goryachev AB, Toh D-J, Wee KB, Lee T, Zhang H-B, Zhang L-H (2005) ''Transition to Quorum Sensing in an Agrobacterium Population: A Stochastic Model.'' PLoS Comput Biol 1(4): e37.]</ref> <ref name="Groisman2005"> [http://www.nature.com/nmeth/journal/v2/n9/pdf/nmeth784.pdf A. Groisman, C. Lobo, H. Cho, J K. Campbell, Y. S. Dufour, A. M Stevens & A. Levchenko. ''A microfluidic chemostat for experiments with bacterial and yeast cells.'' Nature Methods 2005(2), p. 685 - 689] </ref> |
| − | + | |<math> min^{-1} </math> | |
| − | | | + | |<math>8.3 \cdot 10^{-2} s^{-1}</math><ref name="Mehra2008"></ref><ref name="Chatterjee2011"></ref> |
| − | + | ||
| − | | | + | (Range tested: <math>0-0.4 s^{-1}</math>) |
| + | |||
| + | (Bistability range: <math>0.02-0.097 s^{-1}</math><ref name="Mehra2008"></ref> | ||
| + | |||
| + | and <math>8.3 \cdot 10^{-6}-4.2 s^{-1}</math><ref name="Chatterjee2011"></ref>) | ||
| + | | Kaplan et al. conducted experiments on how AHL exits and enters the cell in ''Vibrio fischeri'' and reported that the equimolar internal and external concentrations were established within <math>20 s</math> (estimated rate ~<math>3 min^{-1}</math>), suggesting a very fast diffusion rate. If the signal concentration in the medium is sufficiently large, the time for intracellular concentration to reach equilibrium with the extracellular, is <math>~\frac{1}{D}</math>. Therefore, the rate constant of diffusion can be estimated <math>~ \frac{1}{20}s^{-1}=3 min^{-1}</math>. | ||
| + | [[Image:D-text.png|center|thumb|300px|Kaplan et al. 1985<ref name="Kaplan1985"></ref>]] | ||
| + | |||
| + | Goryachev et al. have published a stochastic model on quorum sensing in ''Agrobacterium tumefaciens'', where they investigated the role of active transport and passive diffusion of the autoinducer AAI in and out of the cell. The values used for their simulations were <math>0.1-0.4 s^{-1} (6-24 min^{-1})</math> for the passive diffusion and <math>0.02-0.6 s^{-1} (1.2-36 min^{-1})</math>. | ||
| + | An estimation within the same range was made by Groisman et al. in a study using a microfluidic chemostat to analyze the cell response to an exogenously added autoinducer, where the reported time span for diffusive exchange of small molecules between different compartments was <math>~40 s</math>. Therefore, the resulting diffusion rate is <math>~ \frac{1}{40}s^{-1}=1.5 min^{-1}</math>. | ||
| + | |||
| + | <div><ul> | ||
| + | <li style="display: inline-block;"> [[Image:D-text2.png|center|thumb|300px|Supplementary material from Goryachev et al. 2005<ref name="Goryachev2005"></ref>-([http://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.0010037#s4 Supporting Information])]]</li> | ||
| + | <li style="display: inline-block;"> [[Image:D-text3.png|center|thumb|350px|Groisman et al. 2005<ref name="Groisman2005"></ref>]] </li> | ||
| + | </ul></div> | ||
| + | |||
| + | Finally, Weber et al. reported some cases of active transport where the diffusion rates are much faster, between <math>~0.01</math> and <math>~0.1 min^{-1})</math>. | ||
| + | [[Image:D-text4.png|center|thumb|350px|Weber et al. 2011<ref name="Weber2011"></ref>]] | ||
|} | |} | ||
Revision as of 04:35, 18 October 2015
The SCB1 protein (C) diffuses from each cell to the environment and back.


Contents
Chemical equation
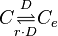
Rate equation
The rate equation is different for the two reactions in order to express the dynamics of the cell density in the environment. Therefore, an additional term 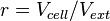 is added in the rate of diffusion from the external environment to the cell. This term represents the additional propensity given to the diffusion of extracellular SCBs by the accumulation of cells in the environment and the consequent increase of cell density. The two rate laws are therefore formed as follows:
is added in the rate of diffusion from the external environment to the cell. This term represents the additional propensity given to the diffusion of extracellular SCBs by the accumulation of cells in the environment and the consequent increase of cell density. The two rate laws are therefore formed as follows:
![r_{out}= D \cdot ([C_{e}]-[C])](/wiki/images/math/b/e/1/be1bfbe3de85b2144bfc73b5cd16a5c2.png)
![r_{in}= r \cdot D \cdot([C]-[C_{e}])](/wiki/images/math/8/4/d/84d9faddb66659ee7bad3500af5bf074.png)
Further information on the coefficient  and the dynamics of cell growth and division is provided here.
and the dynamics of cell growth and division is provided here.
Parameters
The parameter of this reaction is the diffusion rate of C (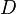 ). In bacteria communication, the autoinducer can either passively diffuse in and out of the cell or, in cases of larger signalling molecules, can be actively transported through the cell membrane. Generally, the diffusion rates when driven by active processes are four orders of magnitude smaller than the passive diffusion rates of small molecules [1]. In this model, active transport is not explored, as SCBs are small molecules that can easily diffuse though the cell membrane. Moreover, transport driven by SCB concentration differences due to increase of the cell density in the environment, is simulated. This means that as cell density grows, more autoinducer molecules enter the cell from the outside. The parameter values were derived from measurements and estimations of different autoinducers' passive diffusion and active transport rates in bacteria.
). In bacteria communication, the autoinducer can either passively diffuse in and out of the cell or, in cases of larger signalling molecules, can be actively transported through the cell membrane. Generally, the diffusion rates when driven by active processes are four orders of magnitude smaller than the passive diffusion rates of small molecules [1]. In this model, active transport is not explored, as SCBs are small molecules that can easily diffuse though the cell membrane. Moreover, transport driven by SCB concentration differences due to increase of the cell density in the environment, is simulated. This means that as cell density grows, more autoinducer molecules enter the cell from the outside. The parameter values were derived from measurements and estimations of different autoinducers' passive diffusion and active transport rates in bacteria.
| Name | Value | Units | Value in previous GBL models [2] [3] | Remarks-Reference |
|---|---|---|---|---|
|
|
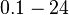 [4] [5] [6] [4] [5] [6]
|
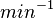
|
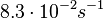 [2][3] [2][3]
(Range tested: (Bistability range: and |
Kaplan et al. conducted experiments on how AHL exits and enters the cell in Vibrio fischeri and reported that the equimolar internal and external concentrations were established within 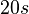 (estimated rate ~ (estimated rate ~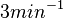 ), suggesting a very fast diffusion rate. If the signal concentration in the medium is sufficiently large, the time for intracellular concentration to reach equilibrium with the extracellular, is ), suggesting a very fast diffusion rate. If the signal concentration in the medium is sufficiently large, the time for intracellular concentration to reach equilibrium with the extracellular, is 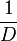 . Therefore, the rate constant of diffusion can be estimated . Therefore, the rate constant of diffusion can be estimated 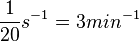 . .
Kaplan et al. 1985[4] Goryachev et al. have published a stochastic model on quorum sensing in Agrobacterium tumefaciens, where they investigated the role of active transport and passive diffusion of the autoinducer AAI in and out of the cell. The values used for their simulations were
Finally, Weber et al. reported some cases of active transport where the diffusion rates are much faster, between  Weber et al. 2011[1] |
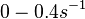 )
)
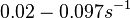
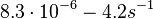
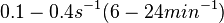 for the passive diffusion and
for the passive diffusion and 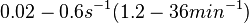 .
An estimation within the same range was made by Groisman et al. in a study using a microfluidic chemostat to analyze the cell response to an exogenously added autoinducer, where the reported time span for diffusive exchange of small molecules between different compartments was
.
An estimation within the same range was made by Groisman et al. in a study using a microfluidic chemostat to analyze the cell response to an exogenously added autoinducer, where the reported time span for diffusive exchange of small molecules between different compartments was 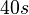 . Therefore, the resulting diffusion rate is
. Therefore, the resulting diffusion rate is 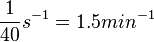 .
.


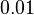 and Failed to parse (Cannot store math image on filesystem.): ~0.1 min^{-1})
.
and Failed to parse (Cannot store math image on filesystem.): ~0.1 min^{-1})
.
{kind=link}
Parameters with uncertainty
The most plausible parameter value for the is decided to be Failed to parse (Cannot store math image on filesystem.): 8 min^{-1}
and the confidence interval Failed to parse (Cannot store math image on filesystem.): 1.5
. This means that the mode of the PDF is 8 and the range where 95% of the values are found is between Failed to parse (Cannot store math image on filesystem.): 4
and Failed to parse (Cannot store math image on filesystem.): 12 min^{-1}
.
The probability distribution for the parameter, adjusted accordingly in order to reflect the above values, is the following:
{kind=link}
The location and scale parameters of the distribution are:
| Parameter | μ | σ |
|---|---|---|
|
|
Failed to parse (Cannot store math image on filesystem.): 2.1267 | Failed to parse (Cannot store math image on filesystem.): 0.2174 |
References
- ↑ 1.0 1.1 Weber M., Buceta J. Noise regulation by quorum sensing in low mRNA copy number systems. BMC Systems Biology 2011, 5:11
- ↑ 2.0 2.1 2.2 S. Mehra, S. Charaniya, E. Takano, and W.-S. Hu. A bistable gene switch for antibiotic biosynthesis: The butyrolactone regulon in streptomyces coelicolor. PLoS ONE, 3(7), 2008.
- ↑ 3.0 3.1 3.2 A. Chatterjee, L. Drews, S. Mehra, E. Takano, Y.N. Kaznessis, and W.-S. Hu. Convergent transcription in the butyrolactone regulon in streptomyces coelicolor confers a bistable genetic switch for antibiotic biosynthesis. PLoS ONE, 6(7), 2011.
- ↑ 4.0 4.1 Kaplan HB, Greenberg EP. Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system. Journal of Bacteriology. 1985;163(3):1210-1214.
- ↑ 5.0 5.1 Goryachev AB, Toh D-J, Wee KB, Lee T, Zhang H-B, Zhang L-H (2005) Transition to Quorum Sensing in an Agrobacterium Population: A Stochastic Model. PLoS Comput Biol 1(4): e37.
- ↑ 6.0 6.1 A. Groisman, C. Lobo, H. Cho, J K. Campbell, Y. S. Dufour, A. M Stevens & A. Levchenko. A microfluidic chemostat for experiments with bacterial and yeast cells. Nature Methods 2005(2), p. 685 - 689