EPAC and PKA-Activation (Agonist)
The EPAC and PKA-Activation is modelled in two different ways. Once to keep as close as possible to Xu et al. (2010)[1] following Michaelis-Menten kinetics (non-modified model) and once as mass action kinetics (modified model), because the activation of those molecules is not due to an enzymatic catalysing but due to binding of cAMP (Kraemer A. et al. 2001)[2].
Binding Reaction
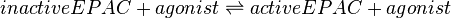
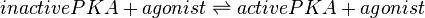
Kinetic Equation
![\frac{k_{cat} \cdot [agonist] \cdot [inactiveEPAC] \cdot (1-\frac{[activeEPAC]}{[inactiveEPAC] \cdot K_{eq}})}{1 + \frac{[inactiveEPAC]}{Km_{inactiveEPAC}} + \frac{[activeEPAC]}{Km_{activeEPAC}}}](/wiki/images/math/b/b/a/bba0627e4f2fa8134fe0c98a0c031d29.png)
![\frac{k_{cat} \cdot [agonist] \cdot [inactivePKA] \cdot (1-\frac{[activePKA]}{[inactivePKA] \cdot K_{eq}})}{1 + \frac{[inactivePKA]}{Km_{inactivePKA}} + \frac{[activePKA]}{Km_{activePKA}}}](/wiki/images/math/e/1/2/e12565a3522e93039695ee14bbe2bc33.png)
final Parameter
Michaelis-Menten:
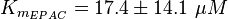
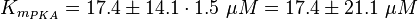
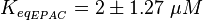
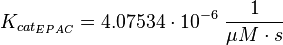
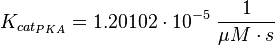
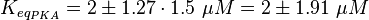
Binding reaction:
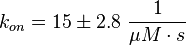
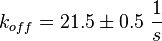
Parameter Annotations
The dissociation constant of cAMP and EPAC is measured, as well as the Km values.(Enserink et al. 2002[3], Takeda et al. 2011[4], Dao et al. 2006[5])
It is assumed that the equilibrium constant is in the range of all tested analoga and therefore the equilibrium constant is calculated by calculating the average and the standard deviation of the both dissociation constants.

Dao et al. (2006)[5]
To calculate the Km the average is used, too. There are three measured values available. Two for the activation by cAMP and one for the activation by 8CPT-2Me-cAMP. Because this reaction is included to increase comparability with Xu et al's model[1] and they used also an analogon for their measurements all three values are averaged. The values are:
KmcAMP(EPAC1) = 30 μM; Enserink et al.(2002)[3]
Km8CPT-2Me-cAMP(EPAC1) = 2.2 μM; Enserink et al. (2002)[3]
KmcAMP(EPAC2) = 20 μM; Takeda et al.(2011)[4]
The error of the Km value of PKA is 1.5 times the one of the value of EPAC.
The kcat values Xu et al. (2010)[1] are used and the averaged relative standard error.
It is shown, that the catalytic domain of EPAC has no influence on the binding ability of EPAC concerning cAMP (Kraemer et al., 2001[2]). Binding experiment between the cAMP-binding domain (cNMPL-EPAC) and cAMP were performed at two different NaCl-level. The dissociation rate differs only slightly, whereas the association rate seems to be dependent on the NaCl-level.
The intracellular sodium concentration in E.coli varies between 5-14 mM (Lo et al. 2006[6]) and can therefore be assumed as less than 50 mM.
Calculation of the kon:
Due to the fact that there are only two values measured, no declaration can be made about the function which describes the relation between the sodium concentration and the association constant. However at higher sodium concentrations a decrease can be observed and this leads to the assumption that at less sodium concentration the association rate might be even higher. It might however be the other way round as well, because maybe the peak of the function is at 50 mM NaCl. We assume that the true value is slightly higher, so the mean is set to 15 1/μMs. To estimate the error the difference of the two measured values is set to 2*σ.
Calculation of the koff: The two values differ only sightly, so we assume that the dissociation rate does not depend on the sodium concentration. Therefore the two values are averaged to get the mean and the error is estimated similar to kon.
The measured values are: koncNMP_L-Epac = 14.4 1/μMs; 50 mM NaCl (Kraemer et al., 2001)[2]
koncNMP_L-Epac= 8.8 1/μMs; 300 mM NaCl (Kraemer et al., 2001)[2]
koffcNMP_L-Epac = 22 s-1; 50 mM NaCl (Kraemer et al., 2001)[2]
koffcNMP_L-Epac = 21 s-1; 300 mM NaCl (Kraemer et al., 2001)[2]
References
- ↑ 1.0 1.1 1.2 Tian-Rui Xu et al. (2010) "Inferring signaling pathway topologies from multiple perturbation measurements of specific biochemical species." Sci Signal. 3(134):ra20. (pmid:20234003)
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 Kraemer A. et al. (2001) "Dynamic interaction of cAMP with the Rap guanine-nucleotide exchange factor Epac1." J Mol Biol 306.5:1167-1177. (pmid:11237625)
- ↑ 3.0 3.1 3.2 Enserik J.M. et al. (2002) "A novel Epac-specific cAMP analogue demonstrates independent regulation of Rap1 and ERK." Nat Cell Biol. 4.11:901-906 (pmid:12402047)
- ↑ 4.0 4.1 Takeda S. et al. (2011) "Systems analysis of GLP-1 receptor signaling in pancreatic β-cells." Am J Pysiol 301.4:C792-C803 (pmid:21734192)
- ↑ 5.0 5.1 Dao K.K. et al. (2006) "Epac1 and cAMP-dependent protein kinase holoenzyme have similar cAMP affinity, but their cAMP domains have distinct structural features and cyclic nucleotide recognition." J Biol Chem 281.30:21500-21511 (pmid:16728394)
- ↑ Lo C., Leake M.C. and Berry R.M. (2006) "Fluorescence measurement of intracellular sodium concentration in single Escherichia coli cells." Biophys J. 90.1:357-365 (pmid:16227503)