Difference between revisions of "Triosephosphate isomerase"
(→Parameters with uncertainty) |
(→Parameters with uncertainty) |
||
| (15 intermediate revisions by the same user not shown) | |||
| Line 8: | Line 8: | ||
<center><math> v = \frac{ V_{mf}\frac{[DHAP]}{K_{DHAP}} - V_{mr}\frac{[Gly3P]}{K_{Gly3P}} }{1 + \frac{[DHAP]}{K_{DHAP}} + \frac{[Gly3P]}{K_{Gly3P}} } </math></center> | <center><math> v = \frac{ V_{mf}\frac{[DHAP]}{K_{DHAP}} - V_{mr}\frac{[Gly3P]}{K_{Gly3P}} }{1 + \frac{[DHAP]}{K_{DHAP}} + \frac{[Gly3P]}{K_{Gly3P}} } </math></center> | ||
| + | |||
| + | Modified rate law considering thermodynamic constant is | ||
| + | <center><math> v = \frac{ V_{mf}\frac{[DHAP]}{K_{DHAP}}\left(1 - \frac{[Gly3P]}{K_{eq}[DHAP]} \right)}{1 + \frac{[DHAP]}{K_{DHAP}} + \frac{[Gly3P]}{K_{Gly3P}} } </math></center> | ||
== Paramters == | == Paramters == | ||
| Line 37: | Line 40: | ||
==Parameters with uncertainty== | ==Parameters with uncertainty== | ||
| − | * The activity is measured in Activity in the reverse reaction in Hernandez (2006) ''et. al.'' <math>V_{mf}</math> is sampled based on Haldane equation <math>K_{eq} = \frac{V_{forward}*K_{product}}{V_{reverse}*K_{substrate}}</math> using the value <math>K_{eq} = | + | * The activity is measured in Activity in the reverse reaction in Hernandez (2006) ''et. al.'' <math>V_{mf}</math> is sampled based on Haldane equation <math>K_{eq} = \frac{V_{forward}*K_{product}}{V_{reverse}*K_{substrate}}</math> using the value <math>K_{eq} = 0.047</math>, <math>Km_{Gly3P}</math> and <math>Km_{DHAP}</math>. <br> |
| + | '''Alternative-1''' the reported fixed point value can be considered with the standard deviation calculated based on the same ratio of <math>V_{mf}</math> which is <math>\approx 31%</math>. This gives the value <math>V_{mf}=6.19 \pm 1.91 </math><math>U\cdot(\text{mg protein})^{-1}</math><br> | ||
| + | '''Alternative-2''' Calculating <math>V_{mf}</math> from <math>V_{mr}</math> based on Haldane equation which gives the value of 2.911 and with the same percent of erro Std. Dev. is 0.90. | ||
{|class="wikitable" | {|class="wikitable" | ||
| Line 47: | Line 52: | ||
|- | |- | ||
|<math>V_{mf}</math> | |<math>V_{mf}</math> | ||
| − | | Sampled based on the Haldane equation. | + | | Sampled based on the Haldane equation.<br> '''Alternative:''' <math>6.19 \pm 1.91</math> or <math>2.911 \pm 0.90</math> conversion gives <math>402 \pm 124.62</math> |
| − | |<math> mM \times min^{-1} </math> | + | |<math>U\cdot(\text{mg protein})^{-1}</math> <br> <math> mM \times min^{-1} </math> |
| | | | ||
|rowspan="5"| | |rowspan="5"| | ||
| Line 67: | Line 72: | ||
|- | |- | ||
|<math>K_{eq}</math> | |<math>K_{eq}</math> | ||
| − | |<math> | + | |<math>0.047 \pm 0.00697 (4)</math><ref name="synder_1975"></ref> |
|mM | |mM | ||
| − | | | + | |<math>K_{eq}(reverse) = 20.9, and so K_{eq}(forward) = \frac{1}{20.9} = 0.047 </math> |
| + | |} | ||
| + | |||
| + | ===Equilibrium constant=== | ||
| + | {| class="wikitable" | ||
| + | ! Equilibrium constant | ||
| + | ! Conditions | ||
| + | ! Source | ||
| + | |- | ||
| + | | 0.045 | ||
| + | | pH=8, T=25°C | ||
| + | | Bergmeyer ''Methods of enzymatic analysis'' page 515<ref name="bermeyer74">Bergmeyer H.U. (1974) ''Methods of enzymatic analysis'', Publisher: Verlag Chemie (vol 1)</ref> | ||
| + | |- | ||
| + | | 0.041 | ||
| + | | pH=7, T=25°C | ||
| + | | Voet et al.<ref name="voet">Voet, D., Voet., J.G. and Pratt, C. W. (1999) Fundamentals of biochemistry, Wiley</ref> from Newshole et al. (1973) <ref name="newshole73">Newshole, E.A. and Stuart, C. (1973) Regulation in Metabolism, Wiley</ref>p 97:<br/> | ||
| + | <math>\Delta G' = 7.9\ kJ.mol^{-1}</math>, <math>Keq = exp(-\frac{\Delta G'}{RT}) = exp(\frac{-7900}{8.31*298.15}) \approx 0.041</math> | ||
| + | |- | ||
| + | | 0.048 | ||
| + | | pH=7, T=25°C | ||
| + | | Lehninger, (1975)<ref name="lehninger75">Lehninger, A.L. (1975) Biochemistry (2nd edn), Worth</ref> p 408:<br/> | ||
| + | <math>\Delta G' = 7.5\ kJ.mol^{-1}</math>, <math>Keq = exp(-\frac{\Delta G'}{RT}) = exp(\frac{-7500}{8.31*298.15}) \approx 0.048</math> | ||
| + | |- | ||
| + | | 0.0475 | ||
| + | | pH=7, T=25°C | ||
| + | | Lehninger, (1975)<ref name="lehninger75">Lehninger, A.L. (1975) Biochemistry (2nd edn), Worth</ref> p 396. | ||
|} | |} | ||
| + | |||
| + | *Taking average of all those values give <math>0.0457 \pm 0.002863</math> | ||
==References== | ==References== | ||
<references/> | <references/> | ||
Latest revision as of 14:23, 28 October 2014
This enzyme rapidly inter-converts the molecules Dihydroxyacetone phosphate (DHAP) and Glyceraldehyde 3-phosphate (Gly3P). Gly3P is removed as soon as it is formed to be used in the next step of glycolysis.
Contents
Chemical equation
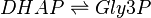
Rate equation
Reversible Michaelis-Menten is used [1]
![v = \frac{ V_{mf}\frac{[DHAP]}{K_{DHAP}} - V_{mr}\frac{[Gly3P]}{K_{Gly3P}} }{1 + \frac{[DHAP]}{K_{DHAP}} + \frac{[Gly3P]}{K_{Gly3P}} }](/wiki/images/math/c/4/3/c43ea2245f63ddca8689602949bdf9f4.png)
Modified rate law considering thermodynamic constant is
![v = \frac{ V_{mf}\frac{[DHAP]}{K_{DHAP}}\left(1 - \frac{[Gly3P]}{K_{eq}[DHAP]} \right)}{1 + \frac{[DHAP]}{K_{DHAP}} + \frac{[Gly3P]}{K_{Gly3P}} }](/wiki/images/math/0/a/2/0a21e55dde0aa7699f6d0a6b766641a7.png)
Paramters
| Parameter | Value | Units | Organism | Remarks |
|---|---|---|---|---|
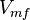
|
5 [1] | 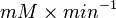
|
Hela cell line | |
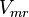
|
42[2] |
| ||
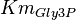
|
0.51[1] | mM | ||
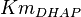
|
1.6[1] | mM |
Parameters with uncertainty
- The activity is measured in Activity in the reverse reaction in Hernandez (2006) et. al. is sampled based on Haldane equation
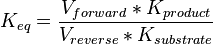 using the value
using the value 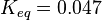 , and .
, and .
Alternative-1 the reported fixed point value can be considered with the standard deviation calculated based on the same ratio of which is 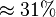 . This gives the value
. This gives the value 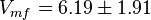
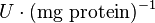
Alternative-2 Calculating from based on Haldane equation which gives the value of 2.911 and with the same percent of erro Std. Dev. is 0.90.
| Parameter | Value | Units | Organism | Remarks |
|---|---|---|---|---|
|
|
Sampled based on the Haldane equation. Alternative: 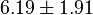 or or 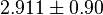 conversion gives conversion gives 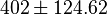
|
|
||
|
|
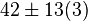 [2] [2]
|
| ||
|
|
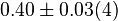 [3] [3]
|
mM | Human liver | |
|
|
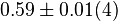 [3] [3]
|
mM | Human liver | |
|
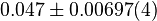 [3] [3]
|
mM | 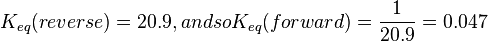
|
Equilibrium constant
| Equilibrium constant | Conditions | Source |
|---|---|---|
| 0.045 | pH=8, T=25°C | Bergmeyer Methods of enzymatic analysis page 515[4] |
| 0.041 | pH=7, T=25°C | Voet et al.[5] from Newshole et al. (1973) [6]p 97:
|
| 0.048 | pH=7, T=25°C | Lehninger, (1975)[7] p 408:
|
| 0.0475 | pH=7, T=25°C | Lehninger, (1975)[7] p 396. |
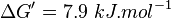 ,
, 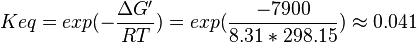
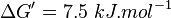 ,
, 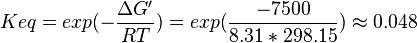
- Taking average of all those values give
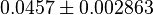
References
- ↑ 1.0 1.1 1.2 1.3 Marín-Hernández A, Gallardo-Pérez JC, Rodríguez-Enríquez S et al (2011) Modeling cancer glycolysis. Biochim Biophys Acta 1807:755–767 (doi) Cite error: Invalid
<ref>tag; name "Hernandez2011" defined multiple times with different content - ↑ 2.0 2.1 Marín-Hernández A , Rodríguez-Enríquez S, Vital-González P A, et al. (2006). Determining and understanding the control of glycolysis in fast-growth tumor cells. Flux control by an over-expressed but strongly product-inhibited hexokinase. FEBS J., 273 , pp. 1975–1988(doi)
- ↑ 3.0 3.1 3.2 Snyder, R.; Lee, E.W. (1975), Triosephosphate isomerase from human and horse liver,Methods Enzymol. 41B, 430-434
- ↑ Bergmeyer H.U. (1974) Methods of enzymatic analysis, Publisher: Verlag Chemie (vol 1)
- ↑ Voet, D., Voet., J.G. and Pratt, C. W. (1999) Fundamentals of biochemistry, Wiley
- ↑ Newshole, E.A. and Stuart, C. (1973) Regulation in Metabolism, Wiley
- ↑ 7.0 7.1 Lehninger, A.L. (1975) Biochemistry (2nd edn), Worth